Archives
Several studies provide evidence of potential
Several studies provide evidence of potential genetic contribution towards AAV (Knight et al., 2008, Monach and Merkel, 2010). The most convincing association has been with the major histocompatibility complex (MHC), especially the locus HLA DPB1*04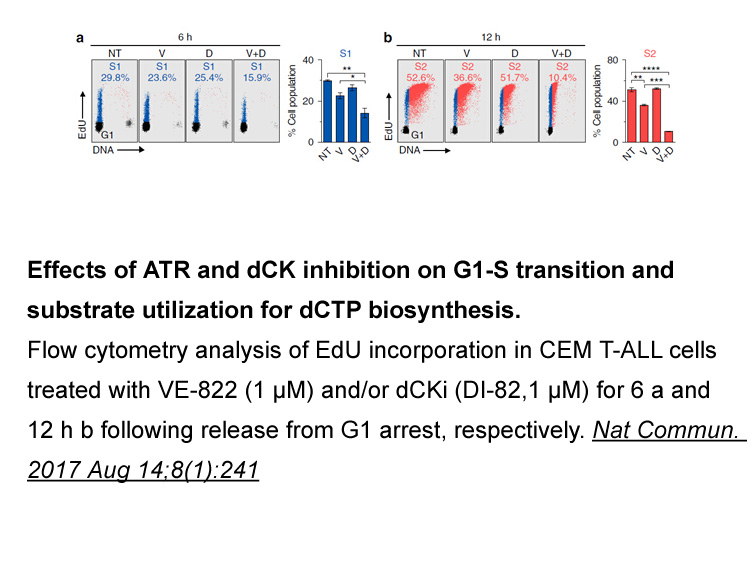 01 (Wieczorek et al., 2010, Jagiello et al., 2004). The other has been suggested between AAV and the rare Z (or null) allele of the serpin A1 gene (SERPINA1) that encodes α1-antitrypsin, a serine proteinase inhibitor for PR3 (Monach and Merkel, 2010; Morris et al., 2011). Both, the HLA-DP and SERPINA1 associations are observed unambiguously in granulomatosis patients with polyangiitis, also positive for PR3-ANCA, but not for MPO-AAV. Thus, the genetic etiology leading to MPO-AAV or MPO-ANCA associated lung injury has remained elusive.
Adropin, a product of the energy homeostasis associated gene (Enho), is a recently identified protein that has been implicated in insulin resistance (Aydin et al., 2013; Kumar et al., 2008) and as a novel regulator of endothelial function (Goetze and Albrethsen, 2014; Lovren et al., 2010). It has been reported that adropin exhibits a potential role to protect endothelium by upregulating endothelial nitric oxide synthase (eNOS) expression through the VEGFR2-PI3K-Akt pathways (Lovren et al., 2010; Ganesh Kumar et al., 2012; Kessenbrock et al., 2009). Our preliminary study found, human MPO-ANCA-related pulmonary hemorrhage displaying adropin-deficiency and target genes mutations. Therefore, we hypothesized that adropin may also exert direct effects on the endothelium in AAV and MPO-ANCA associated lung injury.
01 (Wieczorek et al., 2010, Jagiello et al., 2004). The other has been suggested between AAV and the rare Z (or null) allele of the serpin A1 gene (SERPINA1) that encodes α1-antitrypsin, a serine proteinase inhibitor for PR3 (Monach and Merkel, 2010; Morris et al., 2011). Both, the HLA-DP and SERPINA1 associations are observed unambiguously in granulomatosis patients with polyangiitis, also positive for PR3-ANCA, but not for MPO-AAV. Thus, the genetic etiology leading to MPO-AAV or MPO-ANCA associated lung injury has remained elusive.
Adropin, a product of the energy homeostasis associated gene (Enho), is a recently identified protein that has been implicated in insulin resistance (Aydin et al., 2013; Kumar et al., 2008) and as a novel regulator of endothelial function (Goetze and Albrethsen, 2014; Lovren et al., 2010). It has been reported that adropin exhibits a potential role to protect endothelium by upregulating endothelial nitric oxide synthase (eNOS) expression through the VEGFR2-PI3K-Akt pathways (Lovren et al., 2010; Ganesh Kumar et al., 2012; Kessenbrock et al., 2009). Our preliminary study found, human MPO-ANCA-related pulmonary hemorrhage displaying adropin-deficiency and target genes mutations. Therefore, we hypothesized that adropin may also exert direct effects on the endothelium in AAV and MPO-ANCA associated lung injury.
Materials and Methods
Results
Discussion
ANCA-associated vasculitis (AVV) is a life-threatening condition and patients typically sustain renal and pulmonary injury (Holguin et al., 2008; Lally and Spiera, 2015; Cid, 2012). It has further confirmed that the impaired endothelial function contributes to the development and progression of diverse vascular, inflammatory, and pulmonary hemorrhage (Verma et al., 2003, Dzau et al., 2002). Moreover, endothelium and inflammatory cell infiltration of the neointima is intimately related to oxidative stress (Duckers et al., 2001). In the present study, we showed Enho mutations/adropin-deficiency results in the activation of endothelial Solamargine during neutrophil recruitment and neutrophil-endothelial cell interactions under TNF-α-induced vascular inflammation to be associated with MPO-ANCA associated lung injury.
We identified the serum concentration of adropin was significantly lower in the patients, and we bring hypothesis whether it is associated with Enho mutations. To our surprise, there were not all patients carried Enho mutations. In order to explore the pathogenesis and occurrence of adropin associated with MPO-ANCA-related pulmonary hemorrhage or fibrosis. A microRNA-mRNA interaction network was constructed and it showed that Enho is mainly regulated by miRNAs which revealed that adropin low expression in wild-type patients may be related to the regulation of micro-RNA. To gain more insight into the genotype-phenotype correlation in adropin-deficiency, we used AdrKO mice to investigate the genotype-proteotype-phenotype correlation in patients with adropin-deficiency.
Enho−/− mice showed MPO-ANCA-related pulmonary hemorrhage typical of vasculitis, including red-blood-cell extravasation, a dense perivascular, and predominantly neutrophilic infiltrate in the vascular or bronchus, developed autoantibodies against endothelial cell (VCAM-1 elevated), and autoimmunity pulmonary injury (CD3+, CD20+, and CD38+ cell infiltrate). Elevated serum levels of MPO, CRP, TNF-α, AECA, ET-1, and OPN were detected in AdrKO mice. Furthermore, our study describes the RNA-seq profiling of the pneumonia from AdrKO mice. 507 genes were identified as vascular or autoimmune target candidates. Some examples are cyclin E and mitogen- and stress-activated protein kinases 1 (MSK1) which confirmed cell cycle G1/S progression, monocyte chemoattractant protein-1 gene (MCP-1) for phagocyte recruitment or inflammation, and atrial natriuretic peptide for vascular tone. As compared to the lean mice, the deficiency of adropin decreased Akt1 (Ser473) phosphorylation and downregulated peNOS expression in the lungs of AdrKO mice. It has been reported that Treg cells expressing Foxp3 control the onset and the development of autoimmune disease (Sakaguchi et al., 1995). So, we hypothesized that adropin-deficiency maybe the driver of Treg cells impairment in patients with MPO-ANCA-related lung disease. In vivo, it showed that the proportion and an absolute number of CD4+Foxp3+ cells were significantly decreased in AdrKO mice and these results provide further support for the hypothesis that adropin is a peptide hormone involved in regulating vascular inflammation and autoimmunity.